обозначим меньшую сторону как х а большую как 6х тогда наш периметр равен
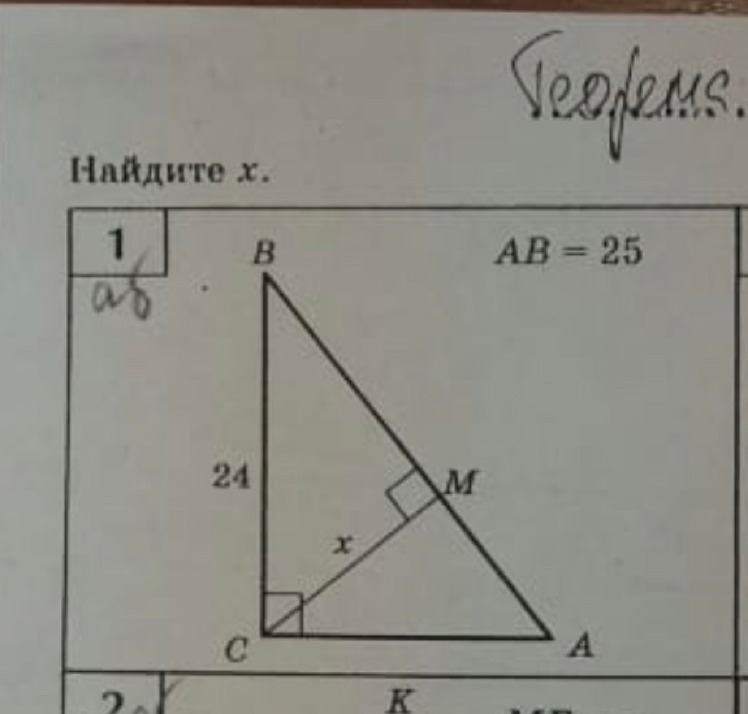
SinA= CB/AB
0,3=9/AB
10/3=AB/9
AB=90/3=30
AC²=AB²-BC²=900-81=819
AC≈28,62
sinB =AC/AB=28,62/30=0,954
cosB=BC/AB=9/30=0,3
tnB=AC/BC=28,62/9=3,18
S=1/2*BC*AC=9*28,62/2=128,79
Ні, не може.
Припустимо, що може бути таки варіант для АВСD чотирикутника<span>, при якому три вершини його А, В і </span>D<span> належать площині </span>α, а вершина С - ні. Проведемо діагоналі АС і ВD<span>. Діагоналі перетинаються в точці О. Оскільки </span>B∈α<span> і </span>D∈α, то ВD належить α<span>, а тому і точка О належить </span>α<span>. Оскільки А</span>∈α<span> і О</span>∈α<span>, то АО належить </span>α<span>. Оскільки точка С належить прямій АО, а пряма АО належить площині </span>α<span>, то і точка С належить площині </span>α. Тому наше припущення не вірне. Не можуть тільки три вершини чотирикутника АВСD <span>належати площині </span>α. Всі чотири лежать в α.
<span>Помимо организмов с типичной клеточной организацией эукариотические клетки) существуют относительно простые, доядерные, или прокариотические, клетки — бактерии и синезеленые, у которых отсутствуют оформленное ядро, окруженное ядерной мембраной, и высокоспециализированные внутриклеточные рганоиды. Особую форму организации живого представляют вирусы и бактериофаги (фаги). Их строение крайне упрощено: они состоят из ДНК (либо РНК) и белкового футляра. Свои функции обмена веществ и размножения вирусы и фаги осуществляют только внутри клеток другого организма: вирусы — внутри клеток растений и животных, </span>
<span>фаги - в бактериальных клетках как паразиты на, генетическом уровне. </span>
<span>К прокариотам относят бактерии и сине-зелёные водоросли (цианеи). Наследственный аппарат прокариот представлен одной кольцевой молекулой ДНК, не образующей связей с белками и содержащей по одной копии каждого гена — гаплоидные организмы. В цитоплазме имеется большое количество мелких рибосом; отсутствуют или слабо выражены внутренние мембраны. Ферменты пластического обмена расположены диффузно. Аппарат Гольджи представлен отдельными пузырьками. Ферментные системы энергетического обмена упорядоченно расположены на внутренней поверхности наружной цитоплазматической мембраны. Снаружи клетка окружена толстой клеточной стенкой. Многие прокариоты способны к спорообразованию в неблагоприятных условиях существования; при этом выделяется небольшой участок цитоплазмы содержащий ДНК, и окружается толстой многослойной капсулой. Процессы метаболизма внутри споры практически прекращаются. Попадая в благоприятные условия, спора преобразуется в активную клеточную форму. Размножение прокариот происходит простым делением надвое. </span>
<span>Средняя величина прокариотических клеток 5 мкм. У них нет никаких внутренних мембран, кроме впячиваний плазматической мембраны. Пласты отсутствуют. Вместо клеточного ядра имеется его эквивалент (нуклеоид), лишенный оболочки и состоящий из одной-единственной молекулы ДНК. Кроме того бактерии могут содержать ДНК в форме крошечных плазмид, сходных с внеядерными ДНК эукариот. В прокариотических клетках, способных к фотосинтезу (сине-зеленые водоросли, зеленые и пурпурные бактерии) имеются различно структурированные крупные впячивания мембраны – тилакоиды, по своей функции соответствующие пластидам эукариот. Эти же тилакоиды или – в бесцветных клетках – более мелкие впячивания мембраны (а иногда даже сама плазматическая мембрана) в функциональном отношении заменяют митохондрии. Другие, сложно дифференцированные впячивания мембраны называют мезасомами; их функция не ясна. </span>
<span>Только некоторые органеллы прокариотической клетки гомологичны соответствующим органеллам эукариот. Для прокариот характерно наличие муреинового мешка – механически прочного элемента клеточной стенки.</span>